
<>Print ![]() PDF
PDF

Самец и самка чечевичника на кормушке в городе.
В продолжение темы, как соотносятся прямые приспособления и изменения под действием отбора
Красивый пример амальгамы прямого приспособления и наследуемых изменений, чей общий фенотипический результат закрепляется отбором (включая устойчивую реализацию в онтогенезе) дают «городские» популяции мексиканского чечевичника Haemorhous mexicanus. У самых разных видов певчих птиц урбанизация сопрягается с ростом частоты вокализаций, составляющих песню, чтобы их «вывести» из полосы техногенного шума, мешающего потенциальным партнёрам распознавать её видоспецифическую структуру (Gil, Brumm, 2014; Halfwerk, Slabbekoorn, 20141). У так называемых «толстоклювых» воробьиных эта частота определяется формой клювного конуса: на примере мексиканского чечевичника исследовали, меняет ли то и другое урбанизация, анализируемая на градиенте переходов от «сельских» местообитаний к «городским» (Giraudeau et al., 2014).
Оказывается, да, у птиц на более урбанизированных территориях клюв делался тоньше и длинней, что повышало частотный диапазон пения и максимальную частоту складывающих его звуков. Как в примерах описанных выше, здесь поведенческие изменения «тянут за собой» морфологические, причём пошедшие через 10 и более лет после формирования городской популяции, когда она уже показала свою жизнеспособность, растёт численно и изменила свою «естественную историю» (Giraudeau et al., 2014).
Самки чечевичников предпочитают самцов, поющих больше песен, и с более длинными песнями. Поэтому важно, чтобы их видоспецифическое звучание не маскировал городской шум (низкочастотный): здесь могут помочь изменения размера и формы клюва, включая угол его раскрытия. Голосовой тракт, трахея, гортань и клюв при пении работают как резонансная камера, а тонкие изменения размера/формы клюва и морфологии голосового тракта сильно влияют на издаваемые звуки. Обычно более длинный, меньше открывающийся, но более широкий клюв обеспечивает значимо меньшие минимум и максимум частоты пения, а также полосу пропускания.
Авторы изучили связь формы клюва и частотной структуры пения мексиканских чечевичников в зависимости от степени урбанизации территорий, лежащих вокруг г. Финикс (Аризона, США); их ряд образует городской градиент.
Степень урбанизации каждой из них оценивали по плотности населения и семи измерениям, касающимся разных паттернов землепользования в 1 км вокруг места поимки птиц (таблица 1). Для численной характеристики степени урбанизации названные 8 метрик анализировали методом главных компонент (PC). Были выделены три координатных оси, PC1-3, вместе объясняющие >84% изменчивости ландшафта, заселенного чеченвичниками: PC1 — 47% дисперсии, PC2 и PC3 — по 24% и 13%. PC1 сильно и негативно коррелировал с % земель, занятых нативным ландшафтом пустыни (вклад этой оси >94%). PC2 сильно и коррелировал с % земель, занятых культивируемой растительностью (вклад >81%), PC3 с % земель, занятых местной растительностью (вклад >74%) (Giraudeau et al., 2014).
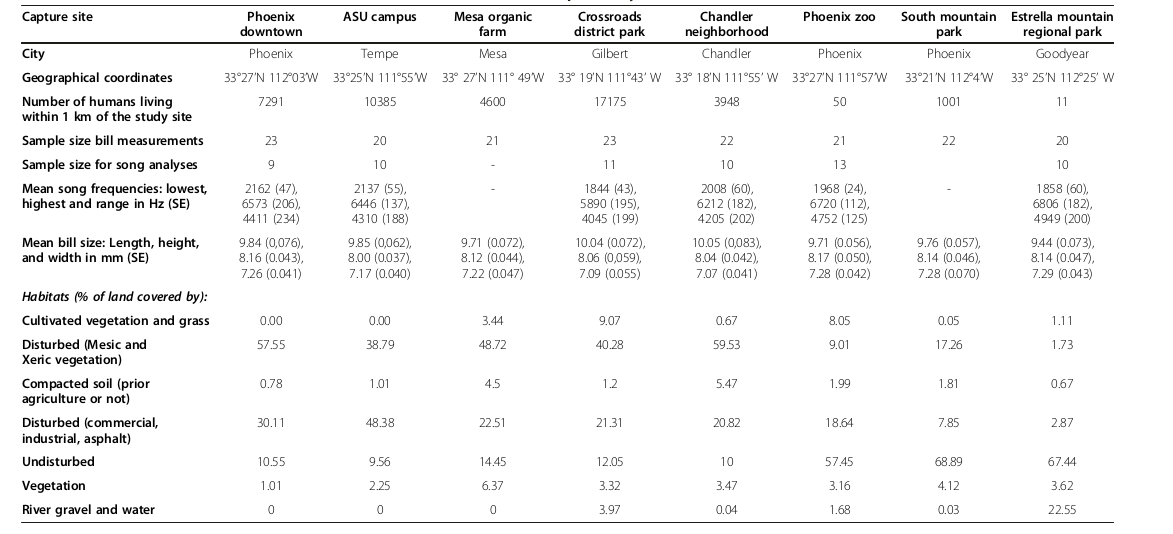 Таблица 1. Изменения формы клюва и песенных характеристик вдоль градиента урбанизации близ г.Финикса: данные, использованные для анализа.
Таблица 1. Изменения формы клюва и песенных характеристик вдоль градиента урбанизации близ г.Финикса: данные, использованные для анализа.
Обозначения. Сверху вниз: населённый пункт, географические координаты, плотность населения в радиусе 1 км, объём выборки для измерений кл.юва, объём выборки для анализ структуры песен, средние характеристики частотной структуры видовой песни: максимум, минимум, диапазон между ними (кГц), структура ландшафта в радиусе 1 км вокруг, % площади, покрытой: возделываемой растительностью, с/хугодья; антропогенно нарушенных (мезо- и ксерофильная растительность); уплотнённой почвой (бывшей ранее в с/хиспользовании или нет); застроенные территории (коммерческая и индустриальная застройка), запечатанные поверхности почвы (асфальт); незастроенные территории; общее покрытие растительности; водное зеркало и песчаные отмели.
Длина цевки и масса тела вьюрков не коррелировали с морфометрией клюва (все P > 0,39). Однако длина цевки (rho = 0,76, P = 0,03) коррелировала с PC2: она удлинена у птиц, отловленных на территориях с большими площадями возделываемой растительности. Форма клюва зависит от степени урбанизации: длина положительно связана с РС1, ширина и высота отрицательно (rho = 0,76, P = 0,03; −0,79, P = 0,02; −0,52, P = 0,18). Чем сильнее пустыня была трансформирована в урбо- или с/хландшафт (т. е. вдоль городского градиента), тем сильней увеличивалась длина клюва чечевичников, а ширина и высота уменьшались (рис.1). Ни один из параметров формы клюва не коррелирует с координатами PC2 и PC3 (все P > 0,49).
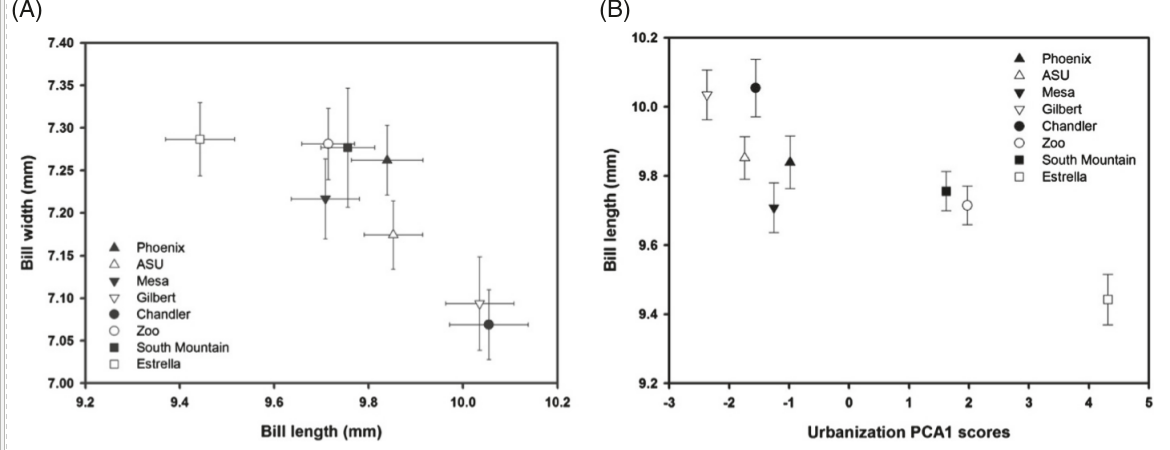 Рисунок 1. Изменения клювного конуса мексиканских чечевичников в зависимости от урбанизованности территории. А. Связи параметров клювного конуса (длина — абсцисса, ширина — ордината, всё в мм) на 8 изученных территориях вокруг Финикса (значки слева внизу). В. Зависимость длины клюва (ордината, мм) от РС1 для тех же 8 участков. Здесь и дальше источник Giraudeau et al., 2014, fig.1-4.
Рисунок 1. Изменения клювного конуса мексиканских чечевичников в зависимости от урбанизованности территории. А. Связи параметров клювного конуса (длина — абсцисса, ширина — ордината, всё в мм) на 8 изученных территориях вокруг Финикса (значки слева внизу). В. Зависимость длины клюва (ордината, мм) от РС1 для тех же 8 участков. Здесь и дальше источник Giraudeau et al., 2014, fig.1-4.
В ряду названных 8 территорий средние ширина и длина клюва связаны отрицательно (rho = -0,86, P = 0,006) при положительной связи ширины с высотой (rho = 0,76, P = 0,03). Последняя не коррелирует с длиной клюва на уровне всей популяции (rho = -0,55, P = 0,16).
Урбанизация и структура песни вьюрков сопряжённо меняются вдоль городского градиента
Содержание
В ряду из 8 территорий средние значения максимальных частот фонового шума отрицательно коррелировали с PC2 (rho = -0,83, P = 0,04), но не с PC1 (rho = 0,03, P = 0,96) или PC3 (rho = 0,60, P = 0,21). Иными словами, сильнее зашумлены территории, меньше покрытые растительностью. В том же ряду самые низкие частоты звуков, образующих видовую песню H.mexicanus, значимо положительно коррелировали с максимумом частоты фонового шума (rho = 0,94, P = 0,005; рис.2); никакие другие характеристики песен не с зашумленностью не связаны (все P>0,6). Минимум частоты пения также отрицательно коррелировал с PC2 (rho = -0,94, P = 0,005), тогда как максимум (rho = -0,83, P = 0,04) и ширина диапазона между ними (rho = -0,84, P = 0,04) коррелируют с РС1 положительно. Вьюрки пели на более высоких частотах в местах, менее покрытых растительностью, и снижали максимум частоты/сужали частотный диапазон на более трансформированных участках.
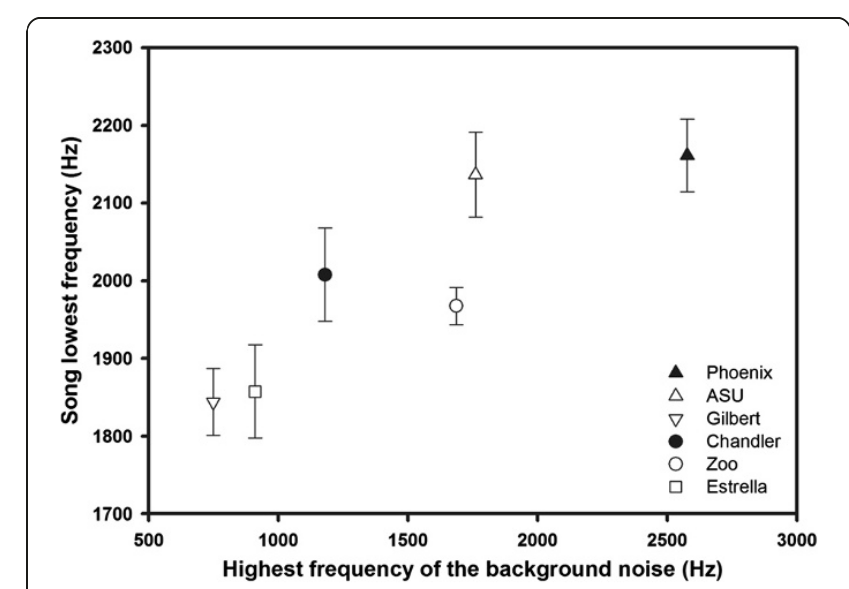 Рисунок 2. Положительная связь минимальной частоты пения и максимальной частоты фонового шума в ряду 6 территорий, для которых можно было собрать данные о структуре песни. Показаны средние ±SE.
Рисунок 2. Положительная связь минимальной частоты пения и максимальной частоты фонового шума в ряду 6 территорий, для которых можно было собрать данные о структуре песни. Показаны средние ±SE.
Сопряжённое варьирование формы клюва и характеристик песни вдоль городского градиента
Различия средней длины и ширины (но не высоты, Р>0,05) клюва вьюрков в ряду территорий значимо связаны с характеристиками их пения. У птиц с более длинными и узкими клювами (это, как помним, связано с трансформированностью ландшафта) ниже максимум песенных частот (rho = -0,94, P = 0,005; рис.3) и уже частотный диапазоном звуков, представленных в песне (rho = -0,94, P = 0,005; рис.4). Самые низкие частоты пения птиц не были связаны с морфологией клюва (все P>0,8). Другими словами, изменения формы клюва, связанные с «городской жизнью», коррелировали с изменением максимума частоты пения и диапазона частот (Girardeau et al., 2014).
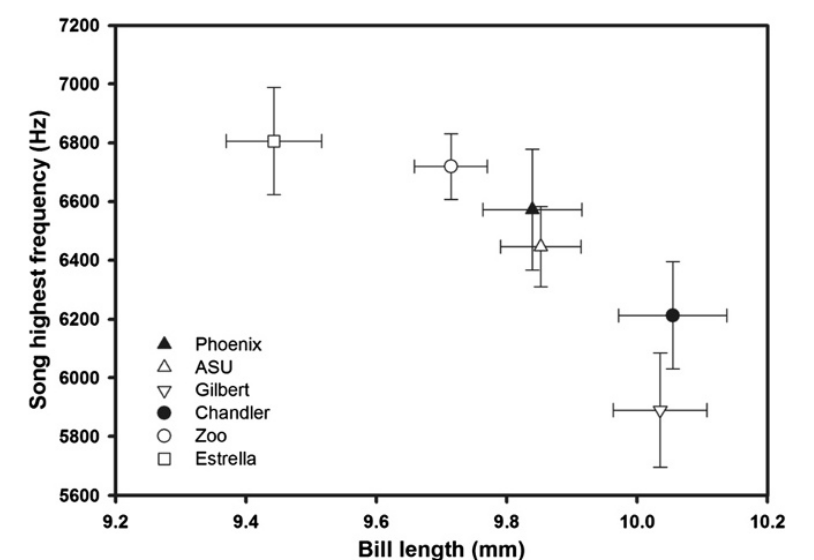 Рисунок 3. Зависимость между максимальными частотами пения (кГц, ордината) и и средней длиной клюва (мм, абсцисса) в ряду 6 территорий (значки слева внизу), для которых получены данные о том и другом признаке (±SE).
Рисунок 3. Зависимость между максимальными частотами пения (кГц, ордината) и и средней длиной клюва (мм, абсцисса) в ряду 6 территорий (значки слева внизу), для которых получены данные о том и другом признаке (±SE).
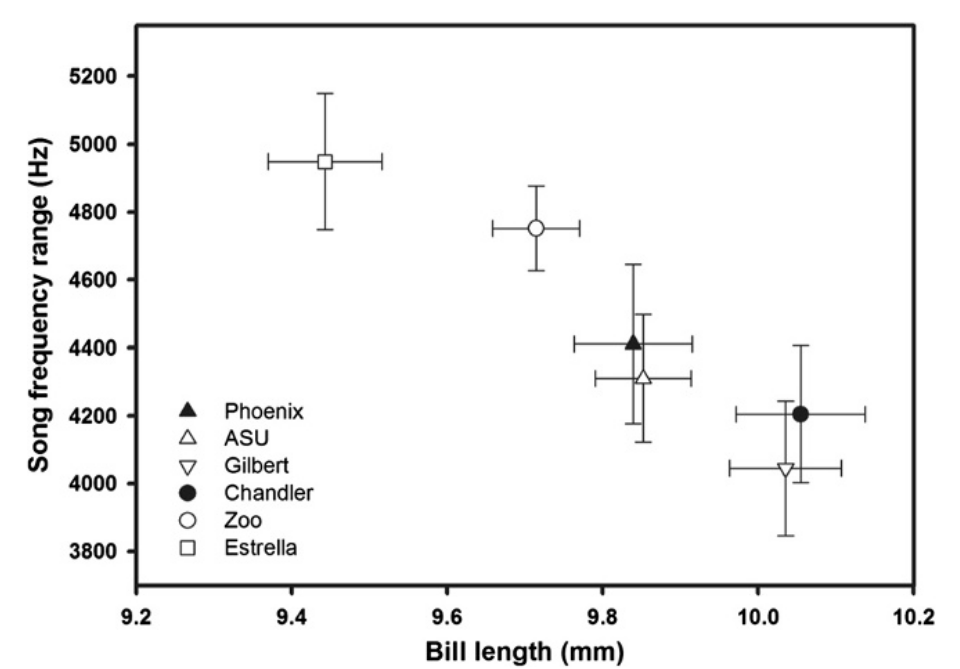 Рисунок 4. Зависимость между шириной частотного диапазона песенных звуков (кГц, ордината) и средней длиной клюва (мм, абсцисса) в ряду 6 территорий (значки слева внизу), для которых получены данные о том и другом признаке (±SE).
Рисунок 4. Зависимость между шириной частотного диапазона песенных звуков (кГц, ордината) и средней длиной клюва (мм, абсцисса) в ряду 6 территорий (значки слева внизу), для которых получены данные о том и другом признаке (±SE).
Обсуждение
Рост степени урбанизации территорий меняет форму клюва чечевичников, он делается длиннее и уже. Обычно различия в форме клюва городских и “сельских” птиц объясняют шаблонно, разным составом кормов. Мол, в городах на кормушках они едят подсолнечник и арахис (более крупные семена с более твёрдой оболочкой), вне города — семена кактусов или сорняков, мелкие, с более тонкой кожурой. Чтобы с первыми справиться, клюв должен быть крупнее, с более сильным укусом. Однако, это не объясняет, почему в Финиксе клювы «городских» чечевичников тоньше и длиннее, чем у «сельских». Самое правдоподобное объяснение — удлинение клюва даёт преимущество в обработке крупных семян на кормушках, а для обработки мелких семян в сельской местности больше годится клюв покороче. Действительно, воробьиные с самыми короткими клювами быстрее всего шелушат самые мелкие семена, более крупноклювые виды быстрее чистят самые крупные (Soobramoney, Perrin, 2007, цит.по: Girardeau et al., 2014).
Точней, изменëнная форма клюва рождается изменением процессов пролиферации и миграции тканей будущего клюва в онтогенезе. Он же в значительной степени регулируется экспрессией костных морфогенетических белков (BMP) на ранних стадиях развития. Если в более урбанизированных районах молодые вьюрки лущат семена с другим уровнем механического воздействия на клюв, то это влияет на выработку BMP, через них — на рост клюва на протяжении целых 2,5 месяцев после вылупления. Недавно Badyaev et al. (2008, цит. по: Girardeau et al., 2014) показали, что разные уровни экспрессии BMP чечевичников в городе и в пустыне могут установиться еще до вылупления. Возможно, [стабилизирующий] отбор рационализировал онтогенез “городских” вьюрков настолько, что поднял уровни экспрессии белков BMP ещё у эмбрионов в яйце, обеспечив формирование у них более длинных и тонких клювов (Girardeau et al., 2014).
Закономерные изменения формы клювного конуса вдоль городского градиента также связаны с изменением структуры видовой песни, прежде всего её частотных характеристик. Более длинный и узкий клюв птиц с более антропогенно трансформированных территорий жёстко связан с уменьшением максимальной частоты пения. Это хорошо согласуется с данными о снижении частоты звучания песен, создаваемых в более длинном голосовом тракте, в том числе за счёт удлинения клюва. Однако минимальная частота пения не снижается, вопреки удлинению клюва с ростом урбанизованности территории, т. е. шире частотный диапазон, в котором звучат песни «более городских» особей. Больше того, максимум уровня фонового шума сильно и положительно коррелирует с минимальной частотой пения чечевичников, что обеспечивает эффективность коммуникации в городе, хотя, будучи более длинноклювыии, в отсутствие необходимости экранировать шум эти птицы могли бы петь сильно ниже (Girardeau et al., 2014).
Видим, что в освоении урбосреды чечевичниками реализон адаптивный компромисс, позволяющий изменяться под одновременным действием двух взаимно противоречивых селективных давлений. Корм из более крупных/жестких семян делает клюв длиннее и тоньше, но не всеми последствиями этого для частотного диапазона пения можно воспользоваться вследствие роста зашумленности в том же ряду территорий. В результате реализованный диапазон у “городских” птиц на 20% уже, чем у пустынных. В эту же сторону “работает” индивидуальная пластичность “выдачи” песенной продукции: столкнувшись с дополнительно созданным шумом или его уменьшением, одни и те же самцы адекватно меняли минимальную частоту пения (Bermúdez-Cuamatzin et al., 2011, цит.по: Girardeau et al., 2014).
Увы, авторам неизвестны каких-либо исследования, показывающие пластичность исполнения на максимальных частотах в ответ на городской шум. В будущих экспериментах следует изучить роль генов, индивидуального развития, обучения и пластичности голосовых путей в формировании разнообразных вокальных характеристик в антропогенной среде.
Далее авторы задаются вопросом: каковы могут быть экологические и эволюционные последствия сопряжённых изменений песни и формы клюва по ходу урбанизации? Особенности песен являются ключевыми показателями качества самцов у многих видов птиц [47]. У домового вьюрка самки продемонстрировали значительные предпочтения в выборе партнера, основанные на характеристиках песен [34]. Следовательно, уменьшение частотного диапазона сигнала в ответ на активность человека может иметь серьезные репродуктивные последствия для мужчин. С другой стороны, гибкость в выборе самок может позволить приемникам использовать альтернативные вокальные компоненты, которые более достоверно отражают мужские качества в новой среде [48],[49]. В соответствии с этой гипотезой, Halfwerk и соавт. (2011, [49]) недавно экспериментально продемонстрировали преимущество самцов больших синиц (Parus major) в передаче сигналов при высокочастотных песнях в шумных условиях, в то время как низкочастотные песни, вероятно, предпочтительнее в естественной, менее шумной среде.
Примечания
Рекомендуем прочесть
Adblock test (Why?)