
<>Print ![]() PDF
PDF

Самец капюшонной вильсонии
Век живи, век учись. Не знал, что у певчих птиц из сем. Древесницевых, Parulidae, белые пятна на хвосте и/или крыльях помогают в ловле добычи, и узнал из работы Ronald R. Mumme (2023). См. каёмки на внешних опахалах рулевых 4-6 у капюшонной вильсонии Setophaga citrina (рис.1). Взмахивая ими, птичка движется в кроне, эти взмахи пугают насекомых, заставляют взлететь и их ловят в угон. Кормление древесницевых эксплуатирует эволюционно консервативную реакцию бегства насекомых; специальные опыты показывают, что закрашивание пятен резко снижает уловистость такого кормления, по сравнению с контролем, где закрашивание лишь изображалось (рис.1 внизу, полевые опыты с S.citrina, а с древесницами р. Myoborus проводили и лабораторные).
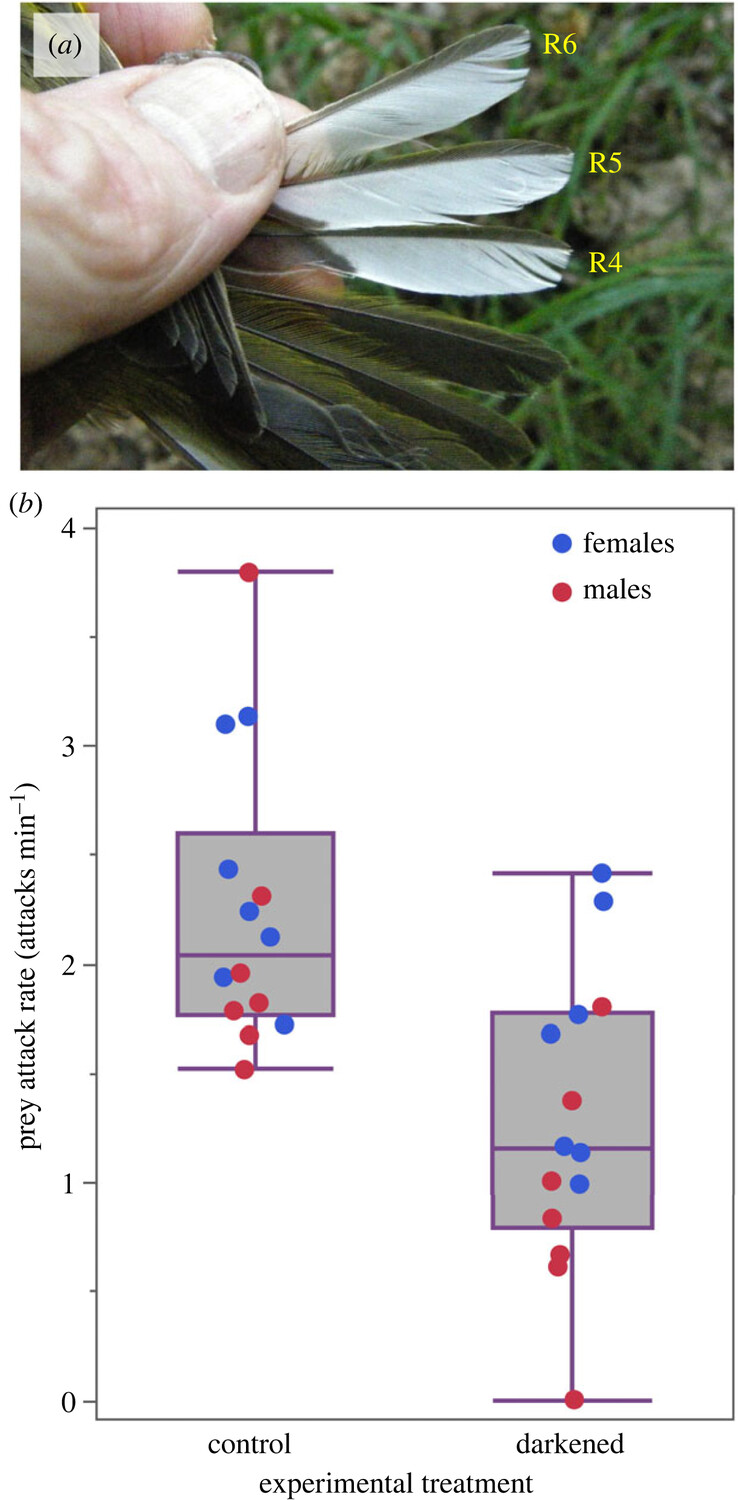
Это верно и для самок и для самцов, хотя у данного вида они на зимовках (т.е.большую часть года, это дальний мигрант) используют разные местообитания: самцы сомкнутые тропические и субтропические леса, самки открытые пространства с кустарниками (скрэб), и ландшафтного разделены. Это работает не везде, скажем, не соответствует фактам попытка объяснить взмахивание хвоста трясогузки использованием этого кормового метода1.
Степень белого на 4-6 парах рулевых у этих птиц варьирует индивидуально, у разных особей их очертания устойчиво сохраняются из года в год и не меняются при позднелетней линьке, с повторяемостью 75%, что заставляет говорить о наследуемости, хотя прямо её не мерили. У данного вида это вообще трудно сделать из-за низкой степени филопатрии и очень высокой доле птенцов, происходящих от внебрачных копуляций.
И вот у капюшонной вильсонии (или древесницы, раньше её относили к ним) Setophaga citrina (рис.3) на данных 14 лет наблюдений за индивидуально меченной популяцией в с.-з. Пенсильвании (Hemlock Hill) в 2010-2023 гг. показан стабилизирующий отбор по величине «ловчих» пятен. В мае-августе здесь картировали все гнездящиеся пары (в разные годы их было 45-70), искали гнезда и фиксировали репродуктивный успех, чтобы связать с ним и с физическим состоянием птиц величину белых кайм на рулях.
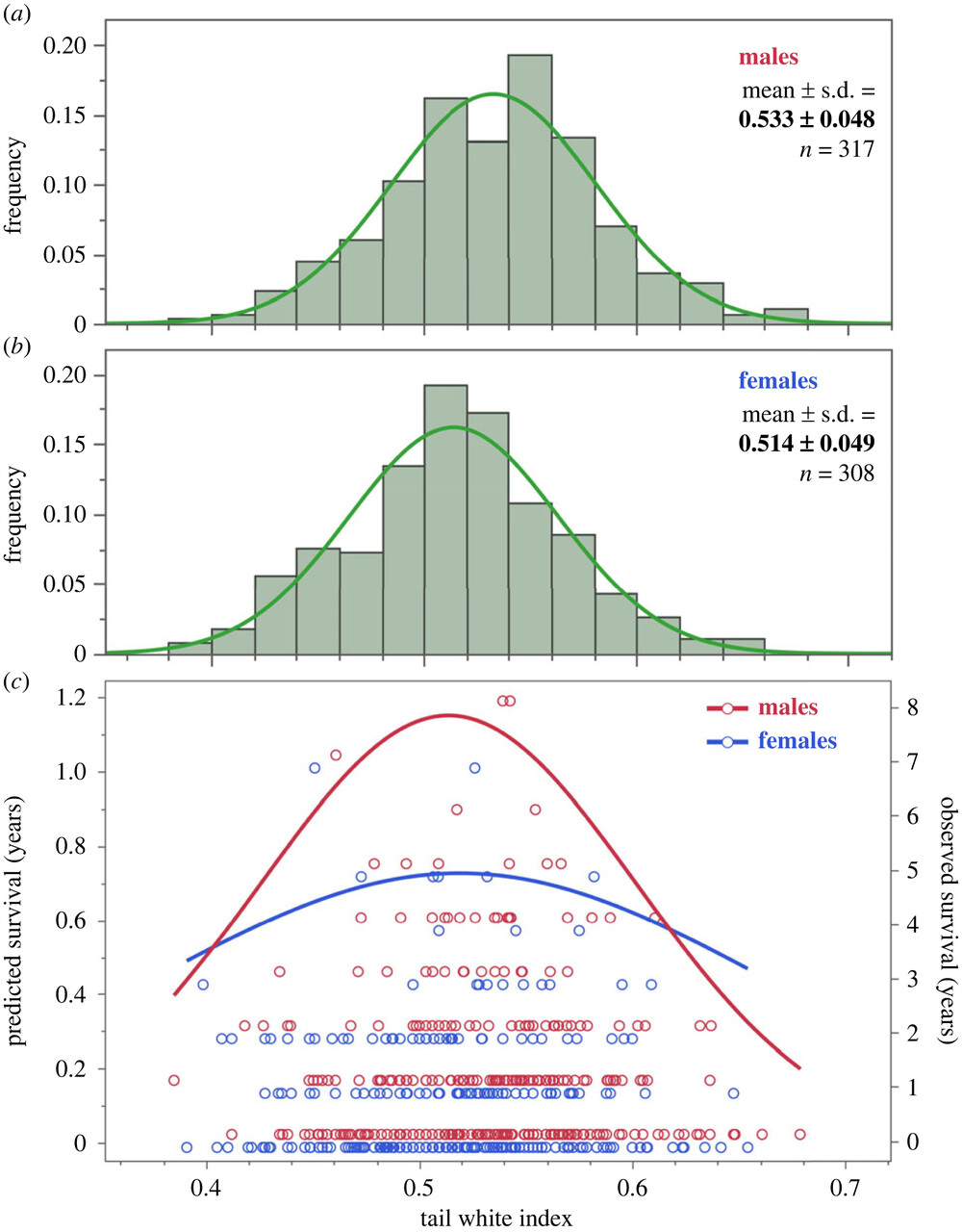
У птиц, пойманных для кольцевания, фиксировали домну крыла, вес, максимальную линейную длину белого на каждом из 3х опахал, а дальше считали индекс развития белого цвета, относя сумму этих 3х промеров к утроенной длине хвоста. Индекс коррелирует с развитием белого на каждом из этих 3х перьев и учитывает колебания длины хвоста. У 28% самцов и 14% самок есть также белое пятнышко на 3ей паре рулевых, но его размер не учитывали, ибо он не коррелирует с величиной других кайм. Для всех птиц, гнездившихся в 2010-2023 гг., и меченных цветными кольцами, посчитана долговременная выживаемость по числу лет присутствия на гнездовании (она была 0, если птица гнездилась лишь 1 сезон).
В среднем это 1,05 (0-8) для самцов (n=317) и 0,78 (0-7, n=308) для самок. Развитость белого на рулях нормально распределена у обоих полов, у самцов она выше, ненамного но значимо (рис.2, верхняя и средняя илл.). У среднего самца каймы примерно на 2 мм длинней, чем у средней самки. Дольше живут особи с пятнами среднего размера, как самцы, так и самки (0,50-0,53, рис.2, нижняя илл.).
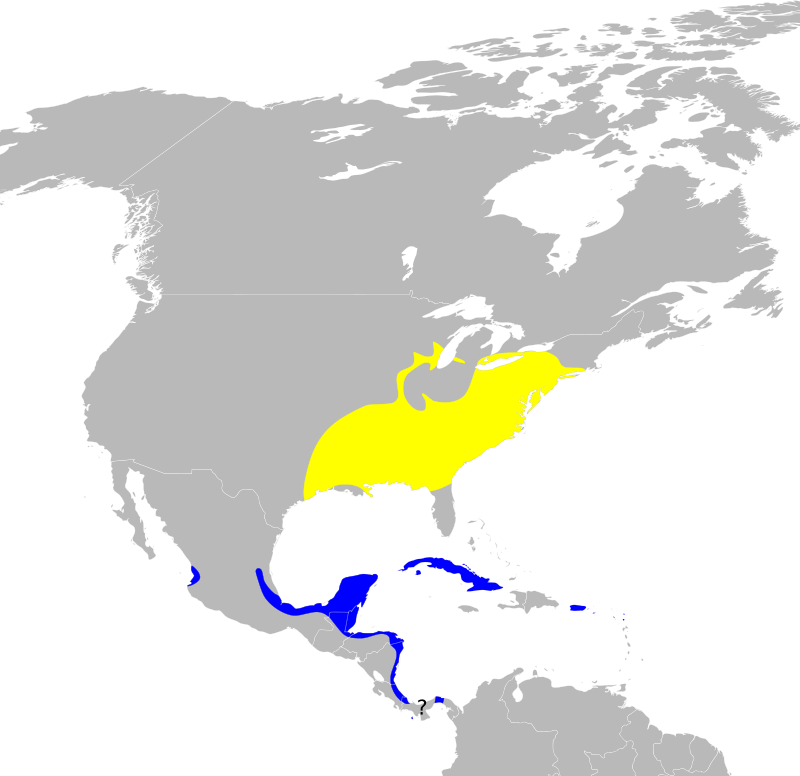
Миграционный путь данного вида: жёлтым — гнездовой, синим — зимовочный ареал
У самцов эффект стабилизирующего отбора выражен сильней, чем у самок, на 5й паре рулевых сильней, чем на 4й и 6й. А вот на репродуктивный успех в данный сезон иди на массу птиц при размножении развитость белого не влияет. Единственный предиктор продукции слетков — число лет пребывания в гнездовой группировке (объясняет 60% изменчивости успеха размножения), массы тела — длина крыла, т. е. общий размер и пол.
Считали, что наблюдаемая выживаемость — случайно распределённая переменная (по Пуассону), её связь с полом и развитием белого на рулях исследовали регрессионным анализом в обобщённой линейной модели со смешанными эффектами (GLMM). Авторы отказались от обычных способов её оценки, вроде модели Кормака — Джолли-Себера, основанной на мечении и повтоном отлове, или модели Кокса, основанной на оценке пропорциональных опасностей, поскольку их интересовали не среднепопуляционные значения этих параметров, а связь наблюдаемой продолжительности жизни [в изученной группировке] и степени развития белого для разных индивидуумов.

Молодая самка на зимовке
Эту связь аппроксимировали сперва линейной, а затем квадратичной зависимостью (табл.1-2), это позволяет «поймать» и исследовать все возможные виды отбора, если они «работают» с ней: движущий (положительный или отрицательный), стабилизирующий, дестабилизирующий или дизруптивный. Пол и развитость белого были фиксированными эффектами в регрессионной модели, год начала наблюдений — случайный эффект. Наблюдаемая долговременная выживаемость недооценивает реальную продолжительность жизни птиц, по двум причинам:
1. Вильсонии, окольцованные в более позднем возрасте, имели впереди меньше лет вероятной жизни, чем окольцованные более молодыми: это корректируется включением года начала исследований как случайного эффекта в модель регрессии.
2. Окончательное исчезновение птиц с территории контролируемой группировки означает не только смерть, но и эмиграцию, которая авторами никак не отслеживалась (она особенно актуальна для самок, их филопатрия ниже, чем у самцов). Однако большая или меньшая развитость белого на рулях вряд ли влияет на вероятность эмиграции и вряд ли «толкает» птиц эмигрировать в какой-то конкретный год жизни: поэтому долговременное пребывание в контролируемой популяции представляется пусть неточной, но и не смещённой оценкой продолжительности жизни в контексте её связи и/или зависимости с развитием белого на рулях, а через него — с эффективностью кормодобывания.
Результаты работы строго доказывают что развитость белого на рулях контролируется стабилизирующим отбором: средние значения развития белого связаны с более долгой жизнью, чем низкие и высокие (у самцов оптимальная степень развития белого значимо выше, чем у самок). Поскольку те, другие и третьи не влияют на успех размножения в каждом из лет, проводимых птицей в данной группировке, её приспособленность прямо пропорциональна продолжительности жизни там. Скорее всего эта связь объясняется вспугивающим эффектом белых пятен определённого размера для тех насекомых, что служат кормом вильсониям так что зависимость между размером пятен и эффективностью кормодобывания описывается оптимальной кривой.
Сниженная приспособленность птиц с большим развитием белого может быть связана также с ещё двумя факторами: белые участки пера легче изнашиваются, чем пигментированные, возможно это ухудшит лётные качества при изнашивании оперения птиц с более белыми рулями или более крупные пятна уже не столько облегчают вспугивание насекомых, сколько делают птицу заметней для хищников. Хотя ни одна из этих двух причин не объясняет пониженной выживаемости птиц с меньшим развитием белого, видимо, здесь действует только меньшая эффективность кормодобывания.
Выявленный авторами стабилизирующий отбор на самцов действует сильней, чем на самок и, главное, поддерживает значимо большую степень развития белого на их рулях, чем у самок. Возможных причин этому две. Первая главная: отбор действует главным образом на зимовке, когда самцы кормятся в тёмных и сомкнутых тропических лесах, самки — в более открытых и светлых зарослях кустарников. В первом случае размер «вспыхивающих» белых пятен для эффективного вспугивания насекомых, обеспечивающего их отлов, должен быть выше, чем во втором, чему есть и экспериментальные доказательства. К тому же растёт количество данных, что доступность пищи на зимовке или, наоборот, смертность из-за её недостатка — главный детерминант продолжительности жизни у дальних мигрантов.
Тем более что у капюшонной вильсонии к тому же размер пятен не влияет ни на репродуктивный успех ни на массу птицы в текущем году (а эти два отклика очевидно связаны с эффективностью кормодобывания в гнездовое время, не на зимовке). Действительно, успех размножения у вильсоний определяется (отрицательно) в первую очередь гибелью гнёзд от хищников и гнездовым паразитизмом коровьих трупиалов Molothrus ater, не эффективностью кормодобывания.

Взрослая самка на гнезде
Вторая причина дополнительная — при ухаживании за самками из чужих пар самцы регулярно расправляют хвосты, так что белые пятна увеличиваются: нужно проверить, имеет ли это сигнальное значение, действительно ли самцы с более крупными пятнами успешней во внебрачных копуляциях. Если да, большая белизна рулевых у самцов связана с ним, тем более что рулевые самцов действительно несколько белей, чем обеспечивало бы наибольшую выживаемость (вершина параболы на рис.2).
Самое интересное и/или важное: выявленный авторами отбор интересен тем, что формирует паттерн окраски вне обычно исследуемого ухаживания или, шире, социальной сигнализации.
Источник Proc. of the Royal Society B. Biol.Sci. 2023. V.290
Рекомендуем прочесть
Adblock test (Why?)